- Postdoctoral Fellow, University of Wisconsin-Madison
- Ph.D., University of Wisconsin-Madison

Scott Michaels
Assistant Vice President for Centers and Core Facilities, IU Research
Professor, Biology
(he/him/his)
Assistant Vice President for Centers and Core Facilities, IU Research
Professor, Biology
(he/him/his)
Myers Hall 359
812-856-0355
Outstanding Junior Faculty Award
NSF CAREER Award
A longstanding interest in my laboratory is the regulation of flowering time. Understanding how undifferentiated cells make developmental decisions is a central challenge in biology. In the model plant Arabidopsis, stem cells in the shoot apical meristem (SAM) give rise to all of the above-ground parts of the plant. Early in development, the SAM gives rise to vegetative structures (e.g., leaves), but later switches to produce the reproductive structures (flowers). The timing of this transition is not predetermined and can be influenced by multiple pathways that integrate both endogenous signals and environmental cues. We are particularly interested in the role of that the floral repressor FLOWERING LOCUS C (FLC) plays in the regulation of flowering. FLC is the major target of the vernalization pathway (the promotion of flowering by cold). Vernalization results in a mitotically stable epigenetic repression of FLC that is mediated by repressive histone modifications, such as H3K27me3. Because of the epigenetic regulation of FLC, our work on flowering time has led to the discovery of a number of epigenetic regulators that play much broader roles in development.
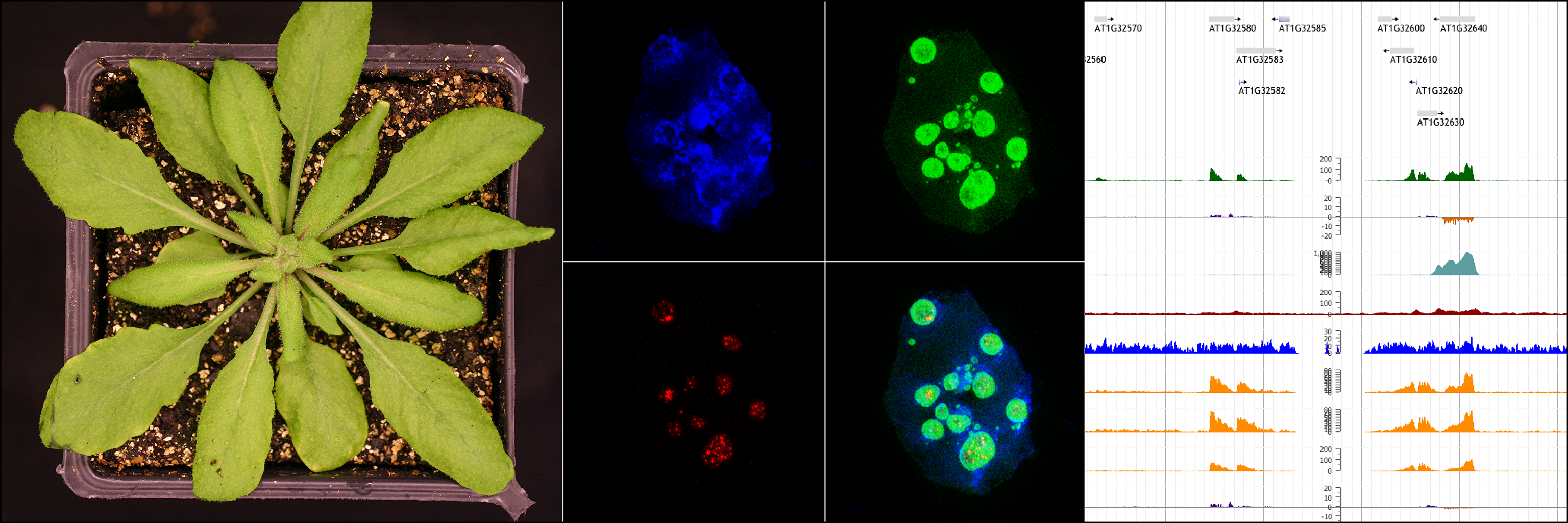
Our current research combines flowering time, gene regulation, epigenetics, and chromatin structure/function. Our laboratory has discovered a previously undescribed class of Arabidopsis transcription factors, BORDER (BDR) proteins, that are providing insight into the relationship between the 3-dimensional arrangement of chromatin and transcription. Changes in gene expression play a central role in development, as well as responses to environmental stimuli/stress. Depending on the situation, hundreds to thousands of genes will show significant changes in expression. A major challenge during these large-scale changes in gene expression is to ensure that changes are restricted to target genes and do not affect the expression of neighboring genes. In animals, CCCTC-binding factor (CTCF) plays a critical role in the formation of chromatin loops and the activity of insulator sequences that separate transcriptional domains. Despite the fact that CTCF homologs are not found in plants, chromatin loops are a significant feature of the Arabidopsis genome, where intragenic “gene loops” often occur between the 5’ and 3’ regions of genes. The function of these gene loops, as well has how they are formed in the absence of CTCF-type insulator proteins, are currently unclear. Our work on BORDER (BDR) proteins are providing insight into the relationship between the 3-dimensional arrangement of chromatin and transcription. BDR proteins interact with RNA polymerase II (Pol II) and are overrepresented in the 5’ and 3’ regions of genes containing gene loops. In bdr mutants, the expression of the loop-containing gene is unaffected, however, the expression of genes that are located downstream on the same DNA strand from the loop-containing gene are downregulated. This suggests a model in which BDR interacts with gene loops to prevent the transcription of upstream genes from interfering with the expression of downstream neighbors. In our future research, will use a range of complementary approaches to investigate the role of BDR proteins in gene loops and transcriptional regulation. These will include using HiC and related techniques to determine the requirement for BDR proteins in the formation and/or function of gene loops, using biochemistry to determine the effect of BDR on Pol II elongation and/or pausing, and using the regulation of flowering time as a system to study the role of BDR proteins in development. Because proteins with BDR-like domain structures occur in plants, animals, and yeast, we believe the knowledge gained in these studies will have broad applications, including human health and food security.
Chromatin, Chromosomes, and Genome Integrity
Developmental Mechanisms and Regulation in Eukaryotic Systems
Plant Molecular Biology
Xianhao Jin; Juzuo Li; Wenqin Lu; Xian Deng; Yanping Wei; Yi Shu; Bo Liu; Zhijian Liu; Yanping Long; Xinlong Zhu; Qili Fei; Qingshun Q. Li; Scott D. Michaels; Xiaofeng Cao; Michael Lenhard; Jixian Zhai (2025) Pre-mRNA processing factors differentially impact coordination between co-transcriptional cleavage and transcription termination (revision under review).
Gen Xu, Jing Lyu, Zhen Wang, Musa Ulutas, Toshihiro Obata, Yufeng Ge, Scott D. Michaels, Sanzhen Liu, James C. Schnable, and Jinliang Yang (2025) Patterns of selection for adaptation to spatial and temporal fluctuating nitrogen availability in maize (in revision).
Zhen Wang, Claudia M Castillo-González, Changjiang Zhao, Chun-Yip Tong, Changhao Li, Songxiao Zhong, Zhiyang Liu, Kaili Xie, Jiaying Zhu, Zhongshou Wu, Xu Peng, Yannick Jacob, Scott D Michaels, Steven E Jacobsen, Xiuren Zhang (2023) “H3.1K27me1 loss confers Arabidopsis resistance to Geminivirus by sequestering DNA repair proteins onto host genome” Nature Communications 14 (1) 7484. PMID: 37980416.
Xuhong Yu, Pascal GP Martin, Yixiang Zhang, Jonathan C Trinidad, Feifei Xu, Jie Huang, Karen E Thum, Ke Li, ShuZhen Zhao, Yangnan Gu, Xingjun Wang, Scott D Michaels (2021) “The BORDER family of negative transcription elongation factors regulates flowering time in Arabidopsis” Current Biology 31(23) 5377-5384. e5.
Matthew T Parker, Katarzyna Knop, Vasiliki Zacharaki, Anna V Sherwood, Daniel Tomé, Xuhong Yu, Pascal Gp Martin, Jim Beynon, Scott D Michaels, Geoffrey J Barton, and Gordon G Simpson. 2021. Widespread premature transcription termination of Arabidopsis thaliana NLR genes by the spen protein FPA. Elife. Apr 27;10:e65537.
Xuhong Yu, Pascal G.P. Martin, and Scott D. Michaels. 2019. “BORDER proteins protect expression of neighboring genes by promoting 3' Pol II pausing in plants.” Nature Commun 10(1) 4359.
Wei Feng, Christopher Hale, Shawn Cokus, Steven E. Jacobsen, and Scott D. Michaels. 2017. Large-scale heterochromatin remodeling linked to over replication-associated DNA damage. PNAS. 114(2) 406-11.
Christopher J. Hale, Magdalena E. Potok, Jennifer Lopez, Truman Do, Ao Liu, Javier Gallego-Bartolome, Scott D. Michaels and Steven E. Jacobsen. 2016. Identification of Multiple Proteins Coupling Transcriptional Gene Silencing to Genome Stability in Arabidopsis thaliana. PLoS Genet. Jun 2;12(6).
Yannick Jacob, Elisa Bergamin, Mark T.A. Donoghue, Vanessa Mongeon, Chantal LeBlanc, Philipp Voigt, Charles J. Underwood, Joseph S. Brunzelle, Scott D. Michaels, Danny Reinberg, Jean-François Couture, and Robert A. Martienssen. 2014. Selective methylation of histone H3 variant H3.1 regulates heterochromatin replication. Science, 343: 1249-1253.
Yannick Jacob, Hume Stroud, Chantal LeBlanc, Suhua Feng, Luting Zhuo, Elena Caro, Christiane Hassel, Crisanto Gutierrez, Scott D. Michaels and Steven E. Jacobsen. 2010. Two histone H3 lysine 27 methyltransferases, ATXR5 and ATXR6, regulate heterochromatic DNA replication. Nature, 466: 987-991.
 The College of Arts
The College of Arts