- Postdoctoral Fellow, Cornell University, 1991-1995
- Ph.D., University of Maryland, 1991

Clay Fuqua
Professor, Biology
Clyde Culbertson Professor, Biology
(he/him/his)
Professor, Biology
Clyde Culbertson Professor, Biology
(he/him/his)
Biology Bldg. 425
812-856-5186
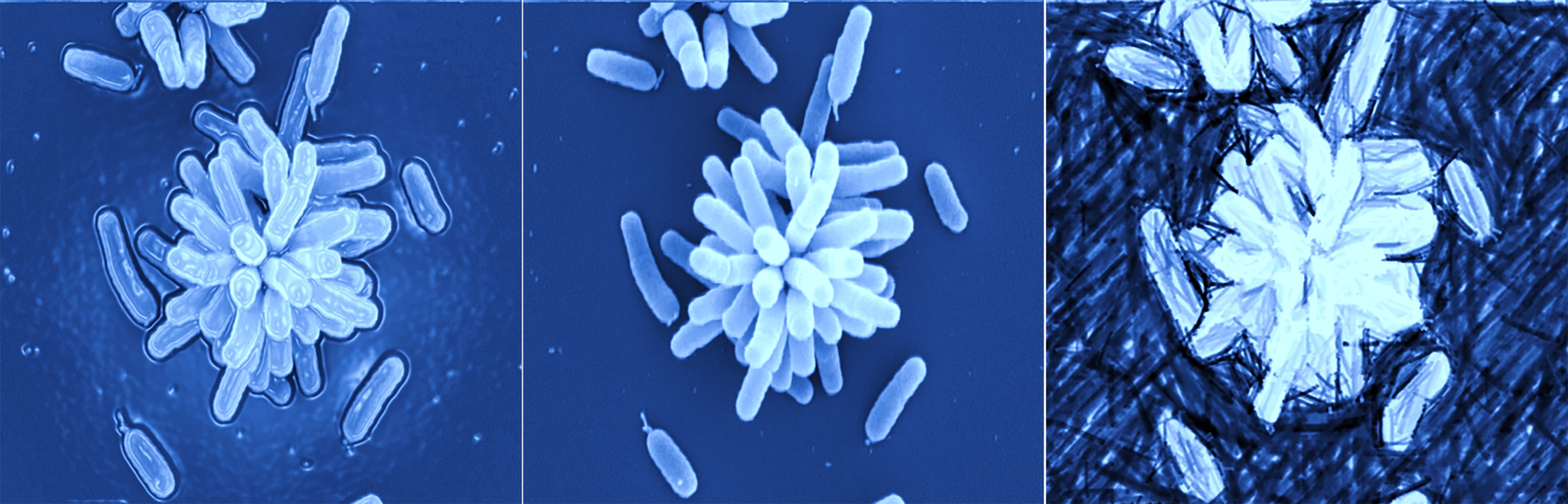
Research projects in the Fuqua laboratory utilize the model Alphaproteobacterium, Agrobacterium tumefaciens, a well-studied bacterial pathogen of plants that causes the disease crown gall. A. tumefaciens is best known for its ability to transfer DNA to plants during pathogenesis, via an interkingdom genetic transfer mechanism that has been extensively utilized to genetically engineer plants. We have discovered that this rod-shaped bacterium exhibits profound asymmetries at the subcellular level that are intimately tied to its cell biology, physiology, genetics and host interactions. One of these asymmetric activities is the production of an adhesive glue at one end of the cell, a structure we call the unipolar polysaccharide (UPP). The UPP is required to cement A. tumefaciens to biotic and abiotic surfaces during formation of the multicellular structures known as biofilms. A. tumefaciens forms robust biofilms on a variety of surfaces, and production of the UPP is a crucial first step in this process. The UPP adhesive is only produced once the cell contacts the surface, and as such is very tightly regulated. We are studying the mechanism of UPP production, its spatial and temporal regulation, and the complex control networks that coordinate attachment with other aspects of cellular physiology and the bacterial cell cycle. Other surface structures such as flagella and pili are also asymmetrically distributed on the cell surface, reflecting a discrete underlying cellular architecture. Recent work demonstrates that A. tumefaciens cells divide asymmetrically via a budding mechanism, in contrast to the standard paradigm of binary fission. We are interested in how the extensive asymmetric organization of the A. tumefaciens cell, its division, and all the activities that it contains, are coordinated. These basic properties are also shared with a growing range of different bacteria, including pathogens and commensals, and thus our fundamental work is broadly relevant.
We also utilize A. tumefaciens as an ecological and evolutionary model for facultative pathogens; bacteria with complex ecologies, that engage in disease under certain conditions, but also have an extensive host-independent existence. Many of the A. tumefaciens populations in the natural environment live as heterotrophic saprophytes, not engaged in plant disease. How do the environmental reservoirs for A. tumefaciens impact the dynamics of disease, and the propensity of this pathogen for virulence? A. tumefaciens has a complex genome, with multiple chromosomes, and megaplasmids. Virulence functions are carried on the tumor-inducing (Ti) plasmid, a 200 kb, self-transmissible plasmid that is required for pathogenesis. A second large, self-transmissible plasmid, called the At plasmid, ranging from 300-800 kb, encodes many functions that enhance A. tumefaciens survival in the rhizosphere (the soil environment influenced by plants) but is not directly involved in pathogenesis. We are interested in how these plasmids shape and are shaped by the ecology and dynamics of disease for A. tumefaciens, how they interact with each other, and how they impact the fitness and virulence of this facultative pathogen in its diverse environments.
Microbial Cell Biology and Environmental Responses
Microbial Interactions and Pathogenesis
Greenwich, J.L., J.L. Eagan, N. Feirer, K. Boswinkle, G. Minasov, L. Shuvalova, N.L. Inniss, J. Raghavaiah, A.K. Ghosh, K.J.F. Satchell, K.D. Allen and C. Fuqua. 2024. Control of biofilm formation by an Agrobacterium tumefaciens pterin-binding periplasmic protein conserved among diverse Proteobacteria. Proc. Natl. Acad. Sci., USA, 121:e2319903121.
Greenwich, J.L., D. Fleming, E. Banin, S. Häussler, B.V. Kjellerup, K. Sauer, K.L. Visick and C. Fuqua. 2023. Conference review: The biofilm community resurfaces: new findings and post-pandemic progress. J. Bacteriol. 205(10):e0016623.
Brown, P.B., J. Chang and C. Fuqua. 2023. Agrobacterium tumefaciens: a transformative agent for fundamental insights into host-microbe interactions, genome biology, chemical signaling and cell biology J. Bacteriol. 9:e0000523.
Liao, Q., Z. Ren, E.E. Wiesler, C. Fuqua and X. Wang. 2022. A dicentric bacterial chromosome requires XerC/D site-specific recombinases for resolution. Curr. Biol. 32:3609-3618.e7, doi:https://doi.org/10.1016/j.cub.2022.06.050; PMCID: PMC9239152.
Onyeziri, M.C., G.G. Hardy, R. Natarajan, J. Xu, I. P. Reynolds, J. Kim, P.M. Merritt, T. Danhorn, M.E. Hibbing, A.J. Weisberg, J.H. Chang and C. Fuqua. 2022. Dual adhesive unipolar polysaccharides synthesized by overlapping biosynthetic pathways in Agrobacterium tumefaciens. Molec. Microbiol., 117, 1023–1047.
Ren, Z., Q. Liao, X. Karaboja, I.S. Barton, E.G. Schantz, A. Mejia-Santana, C. Fuqua, X. Wang. 2022. Conformation and dynamic interactions of the multipartite genome in Agrobacterium tumefaciens. Proc. Natl. Acad. Sci., USA. 119:e2115854119.
Alakavuklar, M.A., B.C Heckel, A.M. Stoner, J.A. Stembel and C. Fuqua. 2021. Motility control through an anti-activation mechanism in Agrobacterium tumefaciens. Molec. Microbiol. 116:1281-97.
Barton, I.S., J.L. Eagan, I.P. Reynolds, P. Nieves-Otero, T.G. Platt and C. Fuqua. 2021. Co-dependent and interdigitated: Dual quorum sensing systems regulate conjugative transfer of the Ti plasmid and the At megaplasmid in Agrobacterium tumefaciens 15955. Front. Microbiol. Sci. 11:1-13.
Labine, M., L DePledge, N. Feirer, J. Greenwich, C. Fuqua, K.D. Allen. 2020. Enzymatic and mutational analysis of the PruA pteridine reductase required for pterin-dependent control of biofilm formation in Agrobacterium tumefaciens. J. Bacteriol. 202:e00098-20.
 The College of Arts
The College of Arts